Nükleotit
| Üzerine bir serinin parçası ⓘ |
| Genetik |
|---|
 |
|
|

Nükleotidler, bir nükleozit ve bir fosfattan oluşan organik moleküllerdir. Nükleik asit polimerlerinin monomerik birimleri olarak görev yaparlar - deoksiribonükleik asit (DNA) ve ribonükleik asit (RNA), her ikisi de Dünya üzerindeki tüm yaşam formlarında temel biyomoleküllerdir. Nükleotidler diyetle elde edilir ve ayrıca karaciğer tarafından yaygın besinlerden sentezlenir. ⓘ
Nükleotidler üç alt birim molekülden oluşur: bir nükleobaz, beş karbonlu bir şeker (riboz veya deoksiriboz) ve bir ila üç fosfattan oluşan bir fosfat grubu. DNA'daki dört nükleobaz guanin, adenin, sitozin ve timindir; RNA'da timin yerine urasil kullanılır. ⓘ
Nükleotidler ayrıca metabolizmada temel, hücresel düzeyde merkezi bir rol oynar. Nükleozit trifosfatlar, adenozin trifosfat (ATP), guanozin trifosfat (GTP), sitidin trifosfat (CTP) ve üridin trifosfat (UTP) şeklinde kimyasal enerji sağlarlar - amino asit, protein ve hücre zarı sentezi, hücre ve hücre parçalarının hareket ettirilmesi (hem hücre içi hem de hücreler arası), hücre bölünmesi gibi enerji gerektiren birçok hücresel işlev için hücre boyunca. Ayrıca nükleotidler hücre sinyalizasyonuna katılır (siklik guanozin monofosfat veya cGMP ve siklik adenozin monofosfat veya cAMP) ve enzimatik reaksiyonların önemli kofaktörlerine dahil edilir (örneğin koenzim A, FAD, FMN, NAD ve NADP+). ⓘ
Deneysel biyokimyada, nükleotidler radyonükleotidler elde etmek için radyonüklidler kullanılarak radyolabelenebilir. ⓘ
5-nükleotidler ayrıca umami tadını arttırmak için gıda katkı maddesi olarak lezzet arttırıcılarda, genellikle maya özütü şeklinde kullanılır. ⓘ

Nükleotitlerin baz kısmı olan nükleobazlar pürinler ve pirimidinler olarak ikiye ayrılır. Adenin ve guanin birer pürin, sitozin, timin ve urasil ise birer pirimidindir. Nükleotitlerin pentoz kısmı riboz veya deoksiriboz'dur. Şeker kısmı deoksiriboz ise nükleotidin adının başına 'deoksi' eklenir. Nükleotidin fosfatsız kısmına nükleozit denir. Nükleozit kısmına bir, iki veya üç fosfat grubu eklenebilir ve bunlara sırasıyla nükleotit monofosfat, difosfat ve trifosfat denir. ⓘ
| Biyokimya ile ilgili bu madde taslak seviyesindedir. Madde içeriğini genişleterek Vikipedi'ye katkı sağlayabilirsiniz. ⓘ |
Yapı

Bir nükleotid üç farklı kimyasal alt birimden oluşur: beş karbonlu bir şeker molekülü, bir nükleobaz - ikisi birlikte nükleozit olarak adlandırılır - ve bir fosfat grubu. Üçü de birleştiğinde bir nükleotid, fosfat grubunu kaç fosfatın oluşturduğuna bağlı olarak "nükleozid monofosfat", "nükleozid difosfat" veya "nükleozid trifosfat" olarak da adlandırılır. ⓘ
Nükleik asitlerde nükleotidler ya bir pürin ya da bir pirimidin bazı, yani azotlu baz olarak da bilinen nükleobaz molekülü içerir ve şeker riboz ise ribonükleotidler, şeker deoksiriboz ise deoksiribonükleotidler olarak adlandırılır. Bireysel fosfat molekülleri, iki komşu nükleotid monomerindeki şeker halkası moleküllerini tekrar tekrar bağlar, böylece bir nükleik asidin nükleotid monomerlerini uzun bir zincir halinde uç uca bağlar. Şeker ve fosfat moleküllerinin bu zincir bağlantıları, tek veya çift sarmal için bir 'omurga' ipliği oluşturur. Herhangi bir sarmalda, zincir bağlantılarının kimyasal yönelimi (yönlülüğü) 5'-uçtan 3'-uca (okunuşu: 5 prime-uçtan 3 prime-uca) uzanır - komşu nükleotidlerdeki şeker molekülleri üzerindeki beş karbon bölgesine atıfta bulunur. Bir çift sarmalda, iki iplik zıt yönlerde yönlendirilir, bu da baz eşleşmesine ve baz çiftleri arasında tamamlayıcılığa izin verir, bunların hepsi DNA'da bulunan kodlanmış bilgilerin çoğaltılması veya kopyalanması için gereklidir. ⓘ
Nükleik asitler, nükleik asitlerin monomer-birimleri olan nükleotidlerden bir araya getirilmiş polimerik makromoleküllerdir. Pürin bazları adenin ve guanin ile pirimidin bazı sitozin hem DNA hem de RNA'da bulunurken, pirimidin bazları timin (DNA'da) ve urasil (RNA'da) sadece birinde bulunur. Adenin, timin ile iki hidrojen bağı ile bir baz çifti oluştururken, guanin sitozin ile üç hidrojen bağı ile çift oluşturur. ⓘ
Nükleik asit polimerlerinin inşası için yapı taşları olmalarının yanı sıra, tekil nükleotidler hücresel enerji depolama ve sağlamada, hücresel sinyalizasyonda, proteinlerin ve diğer sinyal moleküllerinin aktivitesini modüle etmek için kullanılan fosfat gruplarının kaynağı olarak ve genellikle redoks reaksiyonlarını gerçekleştiren enzimatik kofaktörler olarak rol oynarlar. Sinyal siklik nükleotidler, fosfat grubunun aynı şeker molekülüne iki kez bağlanması ve şekerin 5'- ve 3'- hidroksil gruplarının köprülenmesiyle oluşur. Bazı sinyal nükleotidleri, şeker üzerinde farklı konumlara bağlı birden fazla fosfat grubuna sahip olarak standart tek fosfat grubu konfigürasyonundan farklıdır. Nükleotid kofaktörleri, nikotinamid ve flavin de dahil olmak üzere glikozidik bağ yoluyla şekere bağlanan daha geniş bir kimyasal grup yelpazesi içerir ve ikinci durumda riboz şekeri, diğer nükleotidlerde görülen halkayı oluşturmak yerine doğrusaldır. ⓘ


cAMP, hem 5- hem de 3-pozisyonlarına bağlı tek bir fosfata sahip bir siklik nükleotid sinyal molekülü.

pppGpp, hem 5'- hem de 3'-fosfatlı bir nükleotid sinyal molekülü.
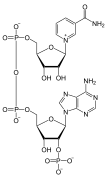
NADP, bir dinükleotid enzimatik kofaktör.

FAD, riboz şekerlerinden birinin halka yerine doğrusal bir konfigürasyon benimsediği bir dinükleotid enzimatik kofaktör. ⓘ




Sentez
Nükleotidler hem in vitro hem de in vivo olarak çeşitli yollarla sentezlenebilir. ⓘ
In vitro olarak, nükleotidlerin laboratuvarda üretimi sırasında koruyucu gruplar kullanılabilir. Saflaştırılmış bir nükleosid, daha sonra doğada bulunmayan analogları elde etmek ve/veya bir oligonükleotid sentezlemek için kullanılabilecek bir fosforamidit oluşturmak üzere korunur. ⓘ
İn vivo olarak, nükleotidler de novo sentezlenebilir veya kurtarma yolları aracılığıyla geri dönüştürülebilir. De novo nükleotid sentezinde kullanılan bileşenler karbonhidrat ve amino asit metabolizmasının biyosentetik öncüllerinden ve amonyak ve karbondioksitten türetilir. Son zamanlarda hücresel bikarbonat metabolizmasının mTORC1 sinyali ile düzenlenebileceği de gösterilmiştir. Karaciğer, dört nükleotidin de novo sentezinin yapıldığı başlıca organdır. Pirimidinlerin ve pürinlerin de novo sentezi iki farklı yol izler. Pirimidinler ilk olarak sitoplazmada aspartat ve karbamoil-fosfattan ortak öncül halka yapısı orotik aside sentezlenir ve bunun üzerine fosforillenmiş bir ribozil birimi kovalent olarak bağlanır. Ancak pürinler ilk olarak halka sentezinin gerçekleştiği şeker şablonundan sentezlenir. Referans olarak, pürin ve pirimidin nükleotidlerinin sentezleri, belirli bir organel içinde değil, hücrenin sitoplazmasındaki çeşitli enzimler tarafından gerçekleştirilir. Nükleotidler, yararlı kısımları yeni nükleotidler oluşturmak üzere sentez reaksiyonlarında yeniden kullanılabilecek şekilde parçalanır. ⓘ
Pirimidin ribonükleotid sentezi

CTP ve UTP pirimidinlerinin sentezi sitoplazmada gerçekleşir ve glutamin ve CO2'den karbamoil fosfat oluşumu ile başlar. Daha sonra, aspartat karbamoiltransferaz, aspartat ve karbamoil fosfat arasında bir yoğunlaşma reaksiyonunu katalize ederek karbamoil aspartik asit oluşturur ve bu da dihidroorotaz tarafından 4,5-dihidroorotik aside siklize edilir. Bu sonuncusu da dihidroorotat oksidaz tarafından orotata dönüştürülür. Net reaksiyon şöyledir:
- (S)-Dihidroorotat + O2 → Orotat + H2O2 ⓘ
Orotat, fosforile edilmiş bir ribozil birimi ile kovalent olarak bağlanır. Riboz ve pirimidin arasındaki kovalent bağlantı, bir pirofosfat içeren riboz biriminin C1 pozisyonunda ve pirimidin halkasının N1 pozisyonunda gerçekleşir. Orotat fosforibosiltransferaz (PRPP transferaz), orotidin monofosfat (OMP) veren net reaksiyonu katalize eder:
- Orotat + 5-Fosfo-α-D-riboz 1-difosfat (PRPP) → Orotidin 5'-fosfat + Pirofosfat ⓘ
Orotidin 5'-monofosfat, üridin monofosfat (UMP) oluşturmak üzere orotidin-5'-fosfat dekarboksilaz tarafından dekarboksile edilir. PRPP transferaz hem ribozilasyon hem de dekarboksilasyon reaksiyonlarını katalize ederek PRPP varlığında orotik asitten UMP oluşturur. UMP'den diğer pirimidin nükleotidleri türetilir. UMP, iki kinaz tarafından ATP ile iki ardışık reaksiyon yoluyla üridin trifosfata (UTP) fosforile edilir. İlk olarak, UDP'den difosfat üretilir ve bu da UTP'ye fosforile edilir. Her iki adım da ATP hidrolizi ile beslenir:
- ATP + UMP → ADP + UDP ⓘ
- UDP + ATP → UTP + ADP ⓘ
CTP daha sonra CTP sentetazın katalitik aktivitesi ile UTP'nin aminasyonu ile oluşur. Glutamin NH3 donörüdür ve reaksiyon ATP hidrolizi ile de beslenir:
- UTP + Glutamin + ATP + H2O → CTP + ADP + Pi ⓘ
Sitidin monofosfat (CMP), sitidin trifosfattan (CTP) iki fosfat kaybıyla türetilir. ⓘ
Pürin ribonükleotid sentezi
Pürin nükleotidlerini oluşturmak için kullanılan atomlar çeşitli kaynaklardan gelir: ⓘ

 |
Pürin halka atomlarının biyosentetik kökenleri N1, Asp'nin amin grubundan kaynaklanır C2 ve C8 formattan kaynaklanır N3 ve N9'a Gln'nin amid grubu katkıda bulunur C4, C5 ve N7 Gly'den türetilmiştir C6 HCO3- (CO2)'den gelir ⓘ |
Bu öncüllerin pürin halkasına dahil edildiği pürin nükleotidlerinin de novo sentezi, hipoksantin bazının nükleotidi olan dallanma noktası ara maddesi IMP'ye giden 10 aşamalı bir yolla ilerler. AMP ve GMP daha sonra bu ara maddeden ayrı, iki aşamalı yollarla sentezlenir. Böylece, pürin parçaları başlangıçta serbest bazlar olarak değil ribonükleotidlerin bir parçası olarak oluşur. ⓘ
IMP sentezinde altı enzim yer alır. Bunlardan üçü çok işlevlidir:
- GART (reaksiyon 2, 3 ve 5)
- PAICS (reaksiyon 6 ve 7)
- ATIC (reaksiyon 9 ve 10) ⓘ
Yol, PRPP oluşumu ile başlar. PRPS1, öncelikle pentoz fosfat yolu tarafından oluşturulan R5P'yi ATP ile reaksiyona sokarak PRPP'ye aktive eden enzimdir. Reaksiyon, bir pirofosforil grubunun ATP'den R5P'nin C1'ine doğrudan aktarılması ve ürünün C1 ile ilgili α konfigürasyonuna sahip olması bakımından sıra dışıdır. Bu reaksiyon aynı zamanda Trp, His ve pirimidin nükleotidlerinin sentez yollarıyla da paylaşılır. Önemli bir metabolik kavşakta bulunan ve çok fazla enerji gerektiren bu reaksiyon yüksek oranda düzenlenir. ⓘ
Pürin nükleotid biyosentezine özgü ilk reaksiyonda PPAT, PRPP'nin pirofosfat grubunun (PPi) glutamin (N), glisin (N&C), aspartat (N), folik asit (C1) veya CO2'den bağışlanan bir amid nitrojeni ile yer değiştirmesini katalize eder. Bu, pürin sentezindeki kararlı adımdır. Reaksiyon, riboz C1 ile ilgili konfigürasyonun ters çevrilmesiyle gerçekleşir, böylece β-5-fosforibosilamin (5-PRA) oluşur ve gelecekteki nükleotidin anomerik formu oluşturulur. ⓘ
Daha sonra, ATP hidrolizi ile beslenen bir glisin dahil edilir ve karboksil grubu daha önce eklenen NH2'ye bir amin bağı oluşturur. Folik asit koenzimi N10-formil-THF'den bir karbon birimi daha sonra sübstitüe glisinin amino grubuna eklenir ve ardından imidazol halkası kapanır. Daha sonra, glutaminden glisin biriminin ilk karbonuna ikinci bir NH2 grubu aktarılır. Glisin ünitesinin ikinci karbonunun karboksilasyonu eşzamanlı olarak eklenir. Bu yeni karbon, bu kez bir aspartat kalıntısından transfer edilen üçüncü bir NH2 biriminin eklenmesiyle modifiye edilir. Son olarak, formil-THF'den ikinci bir karbon birimi azot grubuna eklenir ve halka kovalent olarak kapatılarak yaygın pürin öncüsü inosin monofosfat (IMP) oluşturulur. ⓘ
İnozin monofosfat iki adımda adenozin monofosfata dönüştürülür. İlk olarak, GTP hidrolizi adenilosüksinat sentaz tarafından IMP'ye aspartat eklenmesini sağlar, karbonil oksijeni bir nitrojenle yer değiştirir ve ara ürün adenilosüksinatı oluşturur. Fumarat daha sonra parçalanarak adenozin monofosfat oluşturur. Bu adım adenilosüksinat liyaz tarafından katalize edilir. ⓘ
İnozin monofosfat, ksantilat oluşturan IMP'nin oksidasyonu ve ardından C2'ye bir amino grubunun eklenmesi ile guanozin monofosfata dönüştürülür. NAD+ oksidasyon reaksiyonunda elektron alıcısıdır. Glutaminden amid grubu transferi ATP hidrolizi ile beslenir. ⓘ
Pirimidin ve pürin degradasyonu
İnsanlarda pirimidin halkaları (C, T, U) tamamen CO2 ve NH3'e (üre atılımı) ayrıştırılabilir. Bununla birlikte, pürin halkaları (G, A) parçalanamaz. Bunun yerine, metabolik olarak inert ürik aside parçalanırlar ve daha sonra vücuttan atılırlar. Ürik asit, GMP guanin ve riboz bazlarına ayrıldığında oluşur. Guanin, ksantine deamine olur ve bu da ürik aside oksitlenir. Bu son reaksiyon geri döndürülemez. Benzer şekilde, AMP IMP'ye deamine olduğunda ürik asit oluşabilir ve riboz birimi hipoksantin oluşturmak üzere çıkarılır. Hipoksantin, ksantine ve son olarak ürik aside oksitlenir. Ürik asit salgılanması yerine, guanin ve IMP, PRPP ve aspartat (NH3 donörü) varlığında geri dönüşüm amacıyla ve nükleik asit sentezi için kullanılabilir. ⓘ
Nükleotidlerin prebiyotik sentezi
Yaşamın kökeni hakkındaki teoriler, makul prebiyotik koşullar altında yaşamın temel yapı taşlarının oluşumuna izin veren kimyasal yolların bilinmesini gerektirir. RNA dünyası hipotezi, ilkel çorbada, RNA'yı oluşturmak için seri olarak birleşen temel moleküller olan serbest yüzen ribonükleotidlerin var olduğunu savunur. RNA gibi karmaşık moleküller, reaktiviteleri fiziko-kimyasal süreçler tarafından yönetilen küçük moleküllerden ortaya çıkmış olmalıdır. RNA, her ikisi de güvenilir bilgi aktarımı ve dolayısıyla Darwinci evrim için gerekli olan pürin ve pirimidin nükleotidlerinden oluşur. Becker ve arkadaşları, pirimidin nükleozitlerinin küçük moleküllerden ve ribozdan yalnızca ıslak-kuru döngülerle nasıl sentezlenebileceğini göstermiştir. Pürin nükleosidleri de benzer bir yolla sentezlenebilir. 5'-mono- ve di-fosfatlar da fosfat içeren minerallerden seçici olarak oluşur ve hem pürin hem de pirimidin bazları ile eş zamanlı poliribonükleotid oluşumuna izin verir. Böylece basit atmosferik veya volkanik moleküllerden başlayarak pürin ve pirimidin RNA yapı taşlarına yönelik bir reaksiyon ağı kurulabilir. ⓘ
Doğal olmayan baz çifti (UBP)
Doğal olmayan bir baz çifti (UBP), DNA'nın laboratuvarda oluşturulan ve doğada bulunmayan tasarlanmış bir alt birimidir (veya nükleobaz). Örnekler arasında d5SICS ve dNaM bulunmaktadır. Hidrofobik nükleobazlar taşıyan bu yapay nükleotidler, DNA'da bir (d5SICS-dNaM) kompleksi veya baz çifti oluşturan iki kaynaşmış aromatik halkaya sahiptir. E. coli, UBP'ler içeren bir plazmidi birden fazla nesil boyunca çoğaltmaya teşvik edilmiştir. Bu, genişletilmiş bir genetik kodu sonraki nesillere aktaran canlı bir organizmanın bilinen ilk örneğidir. ⓘ
Sentetik nükleotidlerin tıbbi uygulamaları
Çeşitli nükleotid türevleri hepatit ve HIV'e karşı antiviral olarak kullanılmıştır. Tenofovir disoproksil, Tenofovir alafenamid ve Sofosbuvir hepatite karşı kullanılan NRTI örnekleridir. Mericitabine, Lamivudine, Entecavir ve Telbivudine gibi bazı ilaçlar ise nükleosidlerdir, ancak fosforilasyon yoluyla biyoaktif nükleotid formlarına metabolize edilirler. ⓘ
Uzunluk birimi
Nükleotid ("nt" olarak kısaltılır), baz çiftinin çift sarmallı nükleik asitler için bir uzunluk birimi olmasına benzer şekilde, tek sarmallı nükleik asitler için ortak bir uzunluk birimidir. ⓘ
Dejenere bazlar için kısaltma kodları
IUPAC nükleotidler için semboller belirlemiştir. Beş (A, G, C, T/U) bazın dışında, özellikle PCR primerlerinin tasarlanmasında sıklıkla dejenere bazlar kullanılır. Bu nükleotid kodları burada listelenmiştir. Bazı primer dizileri, standart olmayan inozin nükleotidini kodlayan "I" karakterini de içerebilir. İnozin tRNA'larda bulunur ve adenin, sitozin veya timin ile eşleşir. Ancak bu karakter aşağıdaki tabloda görünmemektedir çünkü bir dejenerasyonu temsil etmemektedir. İnozin, dejenerasyon "D" ile benzer bir işlev görebilse de, ihtiyaç duyulan her olası eşleşmeyi kapsayan bir nükleotid karışımının temsilinden ziyade gerçek bir nükleotiddir. ⓘ
| Sembol | Açıklama | Temsil edilen üsler ⓘ | ||||
|---|---|---|---|---|---|---|
| A | adenin | A | 1 | |||
| C | sitozin | C | ||||
| G | guanin | G | ||||
| T | timin | T | ||||
| U | urasil | U | ||||
| W | zayıf | A | T | 2 | ||
| S | güçlü | C | G | |||
| M | amino | A | C | |||
| K | KETO | G | T | |||
| R | pürin | A | G | |||
| Y | pirimidin | C | T | |||
| B | A değil (B, A'dan sonra gelir) | C | G | T | 3 | |
| D | C değil (D, C'den sonra gelir) | A | G | T | ||
| H | G değil (H, G'den sonra gelir) | A | C | T | ||
| V | T değil (V, T ve U'dan sonra gelir) | A | C | G | ||
| N | herhangi bir taban (boşluk değil) | A | C | G | T | 4 |