Çiçek


Çiçek, bazen çiçeklenme veya çiçek olarak da bilinir, çiçekli bitkilerde (Angiospermae bölümündeki bitkiler) bulunan üreme yapısıdır. Bir çiçeğin biyolojik işlevi, genellikle sperm ile yumurtaların birleşmesi için bir mekanizma sağlayarak üremeyi kolaylaştırmaktır. Çiçekler, çapraz tozlaşmadan kaynaklanan dış çaprazlamayı (bir popülasyondaki farklı bireylerden gelen sperm ve yumurtaların birleşmesi) kolaylaştırabilir veya kendi kendine tozlaşma meydana geldiğinde kendi kendine tozlaşmaya (aynı çiçekten gelen sperm ve yumurtanın birleşmesi) izin verebilir.
İki tür tozlaşma vardır: kendi kendine tozlaşma ve çapraz tozlaşma. Kendi kendine tozlaşma, anterden gelen polen aynı çiçeğin stigmasına veya aynı bitki üzerindeki başka bir çiçeğe bırakıldığında meydana gelir. Çapraz tozlaşma ise polenlerin bir çiçeğin anterinden aynı türün farklı bir bireyindeki başka bir çiçeğin stigmasına aktarılmasıdır. Kendi kendine tozlaşma, ercik ve karpelin aynı anda olgunlaştığı ve polenin çiçeğin stigmasına inebileceği şekilde konumlandığı çiçeklerde gerçekleşir. Bu tozlaşma, tozlayıcılara besin olarak nektar ve polen sağlamak için bitkinin bir yatırım yapmasını gerektirmez. ⓘ
Bazı çiçekler döllenme olmadan diaspor üretir (partenokarpi). Çiçekler sporangia içerir ve gametofitlerin geliştiği yerdir. Birçok çiçek, polen transferi için vektör olmalarına neden olacak şekilde hayvanlar için çekici olacak şekilde evrimleşmiştir. Döllenmeden sonra, çiçeğin yumurtalığı tohum içeren meyveye dönüşür. ⓘ
Çiçekli bitkilerin üremesini kolaylaştırmanın yanı sıra, çiçekler uzun zamandır insanlar tarafından çevreye güzellik katmak için ve ayrıca romantizm, ritüel, ezoterizm, büyücülük, din, bütünsel tıp ve besin kaynağı olarak beğenilmekte ve kullanılmaktadır. ⓘ






Üreme organlarını dıştan sararak onları dış etkilerden korur. Doğrudan üremeye katılmadığı için 'verimsiz kısım' olarak da adlandırılır. ⓘ
Dikotil bitkilerde çiçek örtüsü 2 kısımdan oluşur: Çanak yapraklar (sepal) ve taç yapraklar (petal). Tohumlu bitkilerin en önemli özelliği, çiçek denen üreme organıyla tohum meydana getirmeleridir. Eşeyli üremeye yarayan ve buna uygun şekilde değişikliğe uğramış yapraklar taşıyan sürgün veya sürgün kısımları çiçek adını alır. Bu kısım sınırlı büyüme gösterir ve çoğunlukla diğer sürgünlerden kesin olarak ayrılır. Döllenme ve ekseriya yavru bitkinin ilk gelişmesi burada olur. ⓘ
Etimoloji
Flower, 17. yüzyılda ayrılmadan önce hem öğütülmüş tahıl hem de bitkilerdeki üreme yapısına atıfta bulunan Orta İngilizce flour kelimesinden gelmektedir. Aslen İtalyan çiçek tanrıçası Flora'nın Latince isminden gelmektedir. İngilizce'de çiçek için kullanılan ilk sözcük blossom'dur, ancak günümüzde yalnızca meyve ağaçlarının çiçeklerini ifade etmektedir. ⓘ
Morfoloji

Bir çiçeğin morfolojisi ya da şekli ve yapısı iki kısımda ele alınabilir: taç yapraklar gibi üreme özelliği olmayan yapılardan oluşan vejetatif kısım; ve üreme özelliği olan ya da eşeyli kısımlar. Basmakalıp bir çiçek, hazne adı verilen kısa bir sap veya eksenin ucuna bağlı dört çeşit yapıdan oluşur. Bu parçaların ya da çiçek organlarının her biri whorl adı verilen bir spiral şeklinde düzenlenmiştir. Dört ana sarmal (çiçeğin tabanından veya en alt düğümden başlayarak yukarıya doğru) kaliks, korolla, androecium ve gynoecium'dur. Kaliks ve korolla birlikte çiçeğin periant adı verilen üreme olmayan kısmını oluşturur ve bazı durumlarda farklılaşmayabilir. Eğer durum böyleyse, tepal olarak tanımlanırlar. ⓘ
Periant
Calyx
Toplu olarak kaliks olarak adlandırılan çanak yapraklar, çiçeğin en dış kıvrımında meydana gelen modifiye yapraklardır. Geniş bir tabana, stomalara, stipüllere ve klorofile sahip olmaları bakımından yaprak gibidirler. Çanak yapraklar genellikle mumsu ve serttir ve gelişirken çiçeği korumak için hızla büyürler. Yaprak döken olabilirler, ancak daha yaygın olarak meyve dağılımına yardımcı olmak için büyürler. Kaliks birbirine kaynaşmışsa gamosepalous olarak adlandırılır. ⓘ
Korolla
Taç yapraklar, korolla ile birlikte, periantın en içteki kıvrımını oluşturan neredeyse veya tamamen lifsiz yaprak benzeri yapılardır. Genellikle narin ve incedirler ve tozlaşmayı teşvik etmek için genellikle renkli, şekilli veya kokuludurlar. Şekil olarak yapraklara benzeseler de, birbirleriyle neredeyse aynı anda oluştukları için stamenlerle daha çok karşılaştırılabilirler, ancak sonraki büyümeleri gecikir. Eğer korolla birbirine kaynaşmışsa buna simpetalous denir. ⓘ
Üreme

Androecium
Androecium ya da stamenler, polen üreten erkek kısımların oluşturduğu kıvrımdır. Stamenler tipik olarak bir filamente veya sapa bağlı iki teka halinde düzenlenmiş dört polen kesesinden oluşan bir anterden oluşur. Anter, mayoz bölünme geçirdikten sonra erkek gametofit olan polene dönüşen mikrosporositleri içerir. Çiçek organları arasında en geniş varyasyonu sergilemelerine rağmen, androecium genellikle sadece bir whorl ile ve sadece nadir durumlarda iki whorl ile sınırlıdır. Stamenler sayı, boyut, şekil, yönelim ve çiçeğe bağlandıkları nokta bakımından çeşitlilik gösterir. ⓘ
Gynoecium
Gynoecium ya da karpeller, çiçeğin en içte bulunan dişi kısmıdır. Her bir karpel polenleri alan bir stigma, sap görevi gören bir stil ve ovülleri içeren bir yumurtalıktan oluşur. Karpeller bir ya da birkaç salkım halinde oluşabilir ve birbirine kaynaştığında genellikle pistil olarak tanımlanır. Yumurtalık içinde, yumurtalar plasenta adı verilen doku parçalarından asılıdır. ⓘ
Ovaryum (yumurtalık), stilus (boyuncuk) ve stigma (tepecik) olmak üzere üç kısımdan oluşur. Ovaryum, dişi üreme organının en alt kısmında bulunur ve tohum taslaklarını taşır. Stilus,dişi üreme organının ortasında bulunur ve stigmada çimlenen polenlerin ovaryuma ulaşmasını sağlar. Stigma ise, tozlaşma ile gelen polenlerin dişi organa tutunmasını ve çimlenmesini sağlar. ⓘ
Varyasyon
Bu düzenleme "tipik" olarak kabul edilse de, bitki türleri çiçek yapısında geniş bir çeşitlilik gösterir. Bir çiçeğin dört ana parçası genellikle işlevlerine göre değil, çiçek tablası üzerindeki konumlarına göre tanımlanır. Birçok çiçekte bazı parçalar eksiktir ya da parçalar başka işlevlere dönüştürülmüş olabilir veya tipik olarak başka bir parça gibi görünebilir. Ranunculaceae gibi bazı familyalarda taç yapraklar büyük ölçüde küçülmüştür ve birçok türde çanak yapraklar renkli ve taç yaprağına benzerdir. Diğer çiçeklerde taç yaprağına benzeyen modifiye stamenler bulunur; Şakayık ve Güllerin çift çiçekleri çoğunlukla petaloid stamenlerdir. ⓘ
Birçok çiçeğin simetrisi vardır. Periant herhangi bir noktadan merkezi eksen boyunca ikiye bölündüğünde ve simetrik yarılar üretildiğinde, çiçeğin aktinomorfik veya düzenli olduğu söylenir. Bu radyal simetriye bir örnektir. Çiçekler ikiye bölündüğünde ve simetrik yarımlar üreten sadece bir çizgi ürettiğinde, çiçeğin düzensiz veya zigomorfik olduğu söylenir. Nadiren de olsa hiç simetriye sahip değillerse asimetrik olarak adlandırılırlar. ⓘ
Çiçekler doğrudan bitkiye tabanlarından bağlı olabilir (sapsız - destekleyici sap veya gövde oldukça azalmıştır veya yoktur). Bir çiçeği ya da çiçek salkımını destekleyen sap ya da gövdeye sapçık denir. Bir sap birden fazla çiçeği destekliyorsa, her bir çiçeği ana eksene bağlayan saplara pedisel denir. Çiçekli bir sapın tepesi, torus veya hazne olarak adlandırılan bir terminal şişlik oluşturur. ⓘ
Türlerin çoğunda tek tek çiçeklerin hem pistilleri hem de stamenleri vardır. Bu çiçekler botanikçiler tarafından mükemmel, biseksüel ya da hermafrodit olarak tanımlanır. Ancak bazı bitki türlerinde çiçekler kusurlu ya da tek cinsiyetlidir: sadece erkek (stamen) ya da dişi (pistil) kısımları vardır. Bu ikinci durumda, eğer bir bitki ya dişi ya da erkek ise, o tür iki evcikli olarak kabul edilir. Ancak, aynı bitki üzerinde tek eşeyli erkek ve dişi çiçekler görüldüğünde, türe tek eşeyli denir. Birçok çiçekte, tozlayıcıları çekmek için kullanılan şekerli bir sıvı üreten bezler olan nektarlar bulunur. Kendi başlarına bir organ olarak kabul edilmezler. ⓘ
Çiçeklenme

Bir eksen üzerinde birden fazla çiçeğe sahip olan türlerde, çiçeklerin toplu kümesine çiçek salkımı denir. Bazı çiçek salkımları, tek bir çiçeğe benzeyen bir formasyonda düzenlenmiş birçok küçük çiçekten oluşur. Bunun en yaygın örneği çok büyük kompozit (Asteraceae) grubunun çoğu üyesidir. Örneğin tek bir papatya veya ayçiçeği bir çiçek değil, bir çiçek başıdır - çok sayıda çiçekten (veya çiçekçikten) oluşan bir çiçek salkımı. Bir çiçek salkımı özelleşmiş saplar ve brakteler olarak bilinen modifiye yapraklar içerebilir. ⓘ
Çiçek diyagramları ve formülleri
Çiçek formülü, bir çiçeğin yapısını belirli harfler, sayılar ve semboller kullanarak temsil etmenin bir yoludur ve çiçek hakkında önemli bilgileri kompakt bir biçimde sunar. Genellikle farklı organların veya belirli türlerin sayı aralıklarını vererek bir taksonu temsil edebilir. Çiçek formülleri 19. yüzyılın başlarında geliştirilmiştir ve o zamandan beri kullanımları azalmıştır. Prenner ve arkadaşları (2010), formülün tanımlayıcı kapasitesini genişletmek için mevcut modelin bir uzantısını tasarlamıştır. Çiçek formüllerinin formatı dünyanın farklı yerlerinde farklılık gösterse de aynı bilgiyi aktarırlar. ⓘ
Bir çiçeğin yapısı çiçek diyagramları aracılığıyla da ifade edilebilir. Şematik diyagramların kullanımı, hem çiçek yapısını hem de evrimini anlamak için bir araç olarak uzun açıklamaların veya karmaşık çizimlerin yerini alabilir. Bu tür diyagramlar, füzyon ve simetrinin varlığı da dahil olmak üzere çeşitli organların göreceli konumları ve yapısal ayrıntılar da dahil olmak üzere çiçeklerin önemli özelliklerini gösterebilir. ⓘ
Gelişim
Bir çiçek, determinate apikal meristemden modifiye edilmiş bir sürgün veya eksen üzerinde gelişir (determinate, eksenin belirli bir boyuta kadar büyüdüğü anlamına gelir). Sıkıştırılmış boğum aralarına sahiptir ve klasik bitki morfolojisinde oldukça değiştirilmiş yapraklar olarak yorumlanan yapıları taşır. Ancak detaylı gelişim çalışmaları, stamenlerin genellikle az ya da çok modifiye edilmiş gövdeler (caulomes) gibi başladığını ve bazı durumlarda dallara bile benzeyebildiğini göstermiştir. Çiçekli bitkilerin androecium gelişimindeki tüm çeşitliliği göz önünde bulundurduğumuzda, modifiye yapraklar (phyllomes), modifiye gövdeler (caulomes) ve modifiye dalcıklar (shoots) arasında bir süreklilik buluyoruz. ⓘ
Geçiş
Çiçeklenmeye geçiş, bir bitkinin yaşam döngüsü boyunca yaptığı en önemli faz değişikliklerinden biridir. Bu geçiş, döllenme ve tohum oluşumu için uygun bir zamanda gerçekleşmeli ve böylece maksimum üreme başarısı sağlanmalıdır. Bu ihtiyaçları karşılamak için bir bitki, bitki hormonlarının seviyelerindeki değişiklikler ve mevsimsel sıcaklık ve fotoperiyot değişiklikleri gibi önemli endojen ve çevresel ipuçlarını yorumlayabilir. Çok yıllık ve iki yıllık bitkilerin çoğu çiçek açmak için vernalizasyona ihtiyaç duyar. Bu sinyallerin moleküler yorumu, Constans, Çiçeklenme Lokusu C ve Çiçeklenme Lokusu T dahil olmak üzere çeşitli genleri içeren florigen olarak bilinen karmaşık bir sinyalin iletilmesi yoluyla gerçekleşir. Florigen, üreme açısından uygun koşullarda yapraklarda üretilir ve bir dizi farklı fizyolojik ve morfolojik değişikliği tetiklemek için tomurcuklarda ve büyüyen uçlarda etki eder.

Geçişin ilk adımı vejetatif gövde primordiasının floral primordiaya dönüşmesidir. Bu, yaprak, tomurcuk ve gövde dokularının hücresel farklılaşmasını üreme organlarında büyüyecek dokuya dönüştürmek için biyokimyasal değişiklikler gerçekleştikçe meydana gelir. Gövde ucunun orta kısmının büyümesi durur veya düzleşir ve yanlar gövde ucunun dış çevresinde kıvrımlı veya spiral bir şekilde çıkıntılar geliştirir. Bu çıkıntılar çanak yapraklara, taç yapraklara, stamenlere ve karpellere dönüşür. Bu süreç bir kez başladığında, çoğu bitkide tersine çevrilemez ve çiçek oluşumu olayının ilk başlangıcı bazı çevresel ipuçlarına bağlı olsa bile gövdeler çiçek geliştirir. ⓘ
Organ gelişimi
ABC modeli, çiçeklerin gelişiminden sorumlu genleri tanımlayan basit bir modeldir. Üç gen aktivitesi, çiçek apikal meristemi içindeki primordia organının gelişimsel kimliklerini belirlemek için kombinatoryal bir şekilde etkileşime girer. Bu gen fonksiyonları A, B ve C olarak adlandırılır. A genleri, apikal meristemin yalnızca dış ve en alt kısmında ifade edilir ve bu da bir çanak yaprak salkımına dönüşür. İkinci kıvrımda hem A hem de B genleri ifade edilir ve taç yaprakların oluşmasına yol açar. Üçüncü kıvrımda, B ve C genleri etkileşime girerek stamenleri oluşturur ve çiçeğin merkezinde sadece C genleri karpelleri meydana getirir. Model, Arabidopsis thaliana ve aslanağzı, Antirrhinum majus'taki anormal çiçekler ve mutasyonlar üzerine yapılan çalışmalara dayanmaktadır. Örneğin, B geninin işlevinde bir kayıp olduğunda, mutant çiçekler her zamanki gibi birinci lülede çanak yapraklarla üretilir, ancak normal taç yaprağı oluşumu yerine ikinci lülede de çanak yapraklar bulunur. Üçüncü halkada B fonksiyonunun eksikliği ancak C fonksiyonunun varlığı dördüncü halkayı taklit eder ve üçüncü halkada da karpel oluşumuna yol açar. ⓘ
Fonksiyon
Bir çiçeğin temel amacı bireyin ve türün üremesidir. Tüm çiçekli bitkiler heterosporludur, yani her bir bitki iki tip spor üretir. Mikrosporlar anterlerin içinde mayoz bölünme ile üretilirken megasporlar yumurtalık içindeki ovüllerin içinde üretilir. Anterler tipik olarak dört mikrosporangia'dan oluşur ve bir ovül bütünleşmiş bir megasporangium'dur. Her iki spor türü de sporangia içinde gametofitlere dönüşür. Tüm heterospor bitkilerde olduğu gibi, gametofitler de sporların içinde gelişir, yani endosporiktirler. ⓘ
Bitki türlerinin çoğunda, bireysel çiçekler hem işlevsel karpellere hem de stamenlere sahiptir. Botanikçiler bu çiçekleri mükemmel ya da biseksüel, türleri de hermafrodit olarak tanımlar. Azınlıktaki bitki türlerinin çiçeklerinde üreme organlarından biri ya da diğeri bulunmaz ve bu türler kusurlu ya da tek cinsiyetli olarak tanımlanır. Eğer bir türün her bir bitkisi her iki cinsiyetten de tek eşeyli çiçeklere sahipse, o zaman tür tek eşeylidir. Alternatif olarak, eğer her bir bitki sadece aynı cinsiyetten tek eşeyli çiçeklere sahipse, o zaman tür dioiktir. ⓘ
Tozlaşma


Çiçeğin birincil amacı üremedir. Çiçekler bitkinin üreme organları olduklarından, polenlerin içinde bulunan spermlerin yumurtalıkta bulunan ovüllerle birleşmesine aracılık ederler. Tozlaşma, polenlerin anterlerden stigmaya doğru hareketidir. Normalde polenler çapraz tozlaşma olarak bilinen bir bitkiden diğerine taşınır, ancak birçok bitki kendi kendine tozlaşabilir. Çapraz tozlaşma tercih edilir çünkü türlerin hayatta kalmasına katkıda bulunan genetik çeşitliliğe izin verir. O halde birçok çiçek tozlaşma için rüzgar, su, hayvanlar ve özellikle de böcekler gibi dış etkenlere bağımlıdır. Ancak kuşlar, yarasalar ve hatta bazı cüce keseli sıçanlar gibi daha büyük hayvanlar da kullanılabilir. Bunu başarmak için çiçekler, polenlerin bir bitkiden aynı türden diğerine aktarılmasını teşvik eden özel tasarımlara sahiptir. Bu sürecin gerçekleşebileceği zaman dilimine (çiçek tamamen genişlediğinde ve işlevsel olduğunda) anthesis denir, dolayısıyla tozlaşma biyolojisi çalışmasına anthecology denir. ⓘ
Çiçekli bitkiler genellikle polenlerinin transferini optimize etmek için evrimsel baskı ile karşı karşıyadır ve bu tipik olarak çiçeklerin morfolojisine ve bitkilerin davranışlarına yansır. Polenler bitkiler arasında bir dizi 'vektör' ya da yöntemle aktarılabilir. Çiçekli bitkilerin yaklaşık %80'i biyotik ya da canlı vektörlerden yararlanır. Diğerleri abiyotik veya cansız vektörler kullanır ve bazı bitkiler birden fazla vektör kullanır, ancak çoğu oldukça uzmanlaşmıştır. ⓘ
Bazıları bu grupların arasında veya dışında yer alsa da, çoğu çiçek aşağıdaki iki geniş tozlaşma yöntemi grubuna ayrılabilir: ⓘ
Biyotik tozlaşma
Biyotik vektörleri kullanan çiçekler, polenleri bir çiçekten diğerine aktarmak için böcekleri, yarasaları, kuşları veya diğer hayvanları çeker ve kullanır. Genellikle şekil olarak özelleşmişlerdir ve polen taneciklerinin tozlaştırıcının cezbedici maddeyi (nektar, polen veya eş gibi) aramak için yere indiğinde vücutlarına aktarılmasını sağlayan bir düzenlemeye sahiptirler. Aynı türün birçok çiçeğinden bu cezbediciyi takip ederken, tozlayıcı polenleri ziyaret ettiği tüm çiçeklerin eşit derecede sivri bir hassasiyetle düzenlenmiş stigmalarına aktarır. Birçok çiçek tozlaşmayı sağlamak için çiçek parçaları arasındaki basit yakınlığa güvenirken, diğerleri tozlaşmayı sağlamak ve kendi kendine tozlaşmayı önlemek için ayrıntılı tasarımlara sahiptir. Çiçekler böcekler (entomophily), kuşlar (ornithophily), yarasalar (chiropterophily), kertenkeleler ve hatta salyangoz ve sümüklü böcekler (malacophilae) gibi hayvanları kullanır. ⓘ
Çekim yöntemleri

Bitkiler bir yerden başka bir yere hareket edemezler, bu nedenle birçok çiçek, dağınık popülasyonlardaki bireyler arasında polen aktarmak için hayvanları çekecek şekilde evrimleşmiştir. En yaygın olarak, entomofil olarak bilinen böcek tozlaşmalı çiçekler; Yunanca'da kelimenin tam anlamıyla "böcek seven". Bu böcekleri çekmek için çiçekler genellikle çeşitli kısımlarında besleyici nektar arayan hayvanları çeken nektar adı verilen bezlere sahiptir. Kuşlar ve arılar, "renkli" çiçekleri aramalarını sağlayan renk görüşüne sahiptir. Bazı çiçekler, tozlayıcılara nektarı nerede arayacaklarını gösteren nektar kılavuzları adı verilen desenlere sahiptir; bunlar yalnızca arılar ve diğer bazı böcekler tarafından görülebilen ultraviyole ışık altında görülebilir. ⓘ
Çiçekler tozlayıcıları kokularıyla da çeker, ancak tüm çiçek kokuları insanlara çekici gelmez; bazı çiçekler çürümüş ete ilgi duyan ve ölü hayvan gibi kokan çiçeklere sahip böcekler tarafından tozlanır. Rafflesia cinsi bitkiler ve titan arum da dahil olmak üzere bunlara genellikle Leş çiçekleri denir. Yarasalar ve güveler de dahil olmak üzere gece ziyaretçileri tarafından tozlaşan çiçekler, tozlaştırıcıları çekmek için muhtemelen kokuya odaklanır ve bu nedenle bu tür çiçeklerin çoğu beyazdır. ⓘ
Çiçekler aynı zamanda şekil olarak da özelleşmiştir ve polen tanelerinin tozlaştırıcının çekicisini aramak için yere indiğinde vücutlarına aktarılmasını sağlayan bir stamen düzenine sahiptir. Diğer çiçekler polen taşıyıcıları çekmek için taklit ya da sahte popülasyon kullanırlar. Örneğin birçok orkide, renk, şekil ve koku bakımından dişi arılara veya eşek arılarına benzeyen çiçekler üretir. Erkekler eş bulmak için bir çiçekten diğerine geçerek çiçekleri tozlaştırır. ⓘ
Tozlayıcı ilişkileri
Birçok çiçeğin bir veya birkaç özel tozlaştırıcı organizma ile yakın ilişkisi vardır. Örneğin birçok çiçek yalnızca belirli bir böcek türünü kendine çeker ve bu nedenle başarılı bir üreme için o böceğe güvenir. Bu yakın ilişki bir birlikte evrim örneğidir, çünkü çiçek ve tozlayıcı uzun bir süre boyunca birbirlerinin ihtiyaçlarını karşılayacak şekilde birlikte gelişmiştir. Ancak bu yakın ilişki, yok olmanın olumsuz etkilerini daha da artırır çünkü böyle bir ilişkide üyelerden birinin yok olması, neredeyse kesinlikle diğer üyenin de yok olması anlamına gelecektir. ⓘ
Abiyotik tozlaşma


Abiyotik ya da cansız vektörler kullanan çiçekler, poleni bir çiçekten diğerine taşımak için rüzgârı ya da daha az yaygın olarak suyu kullanır. Rüzgârla dağılan (anemophilous) türlerde, küçük polen taneleri rüzgârla diğer çiçeklere bazen binlerce kilometre taşınır. Yaygın örnekler arasında otlar, huş ağaçları, fagales takımındaki diğer birçok tür, paçavra otları ve birçok saz bulunur. Tozlayıcıları çekmeye ihtiyaçları yoktur ve bu nedenle büyük, gösterişli veya renkli çiçekler yetiştirme eğiliminde değildirler ve nektarları veya fark edilebilir bir kokuları yoktur. Bu nedenle, bitkiler tipik olarak polen alma şansını artırmak için nispeten büyük, tüylü stigmalara sahip binlerce küçük çiçeğe sahiptir. Entomofil çiçeklerin polenleri genellikle iri, yapışkan ve protein açısından zenginken (tozlayıcılar için bir "ödül" görevi görmesi için), anemofil çiçek polenleri tipik olarak küçük tanelidir, çok hafiftir, pürüzsüzdür ve böcekler için çok az besin değeri vardır. Rüzgarın poleni etkili bir şekilde alıp taşıyabilmesi için, çiçeklerde tipik olarak uzun ince filamentlerin ucuna gevşek bir şekilde tutturulmuş anterler bulunur veya polen rüzgarda hareket eden bir kediciğin etrafında oluşur. Bunun daha nadir biçimleri, tek tek çiçeklerin rüzgar tarafından hareket ettirilmesini (Sarkık) veya daha az yaygın olarak; poleni rüzgara bırakmak için anterlerin patlamasını içerir. ⓘ
Su yoluyla tozlaşma (hidrofil) çok daha nadir bir yöntemdir ve abiyotik olarak tozlaşan çiçeklerin yalnızca yaklaşık %2'sinde görülür. Bunun yaygın örnekleri arasında Calitriche autumnalis, Vallisneria spiralis ve bazı deniz otları sayılabilir. Bu gruptaki çoğu türün paylaştığı bir özellik, polen tanesinin etrafında bir ekzin veya koruyucu tabaka bulunmamasıdır. Paul Knuth 1906'da iki tip hidrofil tozlaşma tanımlamış ve Ernst Schwarzenbach 1944'te buna bir üçüncüsünü eklemiştir. Knuth bu iki gruba Hyphydrogamy ve daha yaygın olan Ephydrogamy isimlerini vermiştir. Hiphidrogamide tozlaşma su yüzeyinin altında gerçekleşir ve bu nedenle polen taneleri tipik olarak negatif kaldırma kuvvetine sahiptir. Bu yöntemi sergileyen deniz bitkilerinde stigmalar genellikle serttir, tatlı su türlerinde ise küçük ve tüylü stigmalar bulunur. Efidrogamide tozlaşma su yüzeyinde gerçekleşir ve bu nedenle polen yüzmeyi sağlamak için düşük bir yoğunluğa sahiptir, ancak birçoğu da sal kullanır ve hidrofobiktir. Deniz çiçekleri yüzen iplik benzeri stigmalara sahiptir ve gelgit için adaptasyonlara sahip olabilirken, tatlı su türleri suda girintiler oluşturur. Schwarzenbach tarafından ortaya konan üçüncü kategori, polenleri taşıma yoluyla suyun üzerine taşıyan çiçeklerdir. Bunlar yüzen bitkilerden (Lemnoideae) staminalı çiçeklere (Vallisneria) kadar uzanır. Bu gruptaki türlerin çoğunda, bazen daha büyük kütleler oluşturan kuru, küresel polenler ve suda çukurlar oluşturan dişi çiçekler bulunur; taşıma yöntemi değişiklik gösterir. ⓘ
Mekanizmalar
Çiçekler iki mekanizma ile tozlaşabilir; çapraz tozlaşma ve kendi kendine tozlaşma. Her birinin avantaj ve dezavantajları olduğu için hiçbir mekanizma diğerinden tartışmasız daha iyi değildir. Bitkiler, habitatlarına ve ekolojik nişlerine bağlı olarak bu mekanizmalardan birini veya her ikisini de kullanır. ⓘ
Çapraz tozlaşma
Çapraz tozlaşma, karpelin aynı türden farklı bir bitkiden gelen polenle tozlaşmasıdır. Diğer bitkiden gelen polenlerin içerdiği spermlerin genetik yapısı farklı olduğundan, bunların birleşimi, eşeyli üreme süreci yoluyla genetik olarak farklı yeni bir bitkiyle sonuçlanacaktır. Her yeni bitki genetik olarak farklı olduğundan, farklı bitkiler fizyolojik ve yapısal adaptasyonlarında çeşitlilik gösterir ve böylece popülasyon bir bütün olarak çevrede meydana gelebilecek olumsuzluklara karşı daha hazırlıklı olur. Dolayısıyla çapraz tozlaşma türlerin hayatta kalmasını arttırır ve bu nedenle genellikle çiçekler tarafından tercih edilir. ⓘ
Kendi kendine tozlaşma
_flowers.jpg)
Kendi kendine tozlaşma, bir çiçeğin karpelinin ya aynı çiçekten ya da aynı bitki üzerindeki başka bir çiçekten gelen polenlerle tozlaşmasıdır ve eşeysiz üreme yoluyla genetik bir klonun oluşmasına yol açar. Bu, tohum üretiminin güvenilirliğini, üretilebilme hızını artırır ve ihtiyaç duyulan enerji miktarını azaltır. Ancak, en önemlisi, genetik çeşitliliği sınırlar. Kendi kendine döllenmenin en uç örneği, ovülün aynı çiçek ya da bitkiden gelen polenle döllenmesi, birçok karahindiba gibi her zaman kendi kendine döllenen çiçeklerde görülür. Bazı çiçekler kendi kendine tozlaşır ve hiç açılmayan çiçeklere sahiptir veya çiçekler açılmadan önce kendi kendine tozlaşır; bu çiçeklere cleistogamous denir; örneğin Viola cinsindeki birçok tür bunu sergiler. Buna karşılık, birçok bitki türünün kendi kendine tozlaşmayı ve dolayısıyla kendi kendine döllenmeyi önleme yolları vardır. Aynı bitki üzerindeki tek cinsiyetli erkek ve dişi çiçekler aynı anda ortaya çıkmayabilir veya olgunlaşmayabilir ya da aynı bitkiden gelen polenler yumurtalarını dölleyemeyebilir. Kendi polenlerine karşı kimyasal engelleri olan ikinci çiçek türleri kendine uyumsuz olarak adlandırılır. Clianthus puniceus'ta (resimde), kendi kendine tozlaşma stratejik olarak bir "sigorta poliçesi" olarak kullanılır. Bir tozlayıcı, bu durumda bir kuş, C. puniceus'u ziyaret ettiğinde stigmatik örtüyü ovar ve kuştan gelen polenin stigmaya girmesine izin verir. Ancak hiçbir tozlayıcı ziyaret etmezse, o zaman stigmatik örtü doğal olarak düşer ve çiçeğin kendi anterlerinin kendi kendine tozlaşma yoluyla çiçeği tozlaştırmasına izin verir. ⓘ
Alerjiler
Polenler, dünya çapında insanların %10 ila 50'sini etkileyen astım ve diğer solunum yolu alerjilerine büyük katkıda bulunmaktadır. İklim değişikliğine bağlı sıcaklık artışları, bitkilerin daha fazla alerjen olan daha fazla polen üretmesi anlamına geldiğinden, bu sayı artıyor gibi görünmektedir. Bununla birlikte, küçük boyutları ve doğal ortamdaki yaygınlığı nedeniyle polenlerden kaçınmak zordur. Alerjiye neden olan polenlerin çoğu otlar, huş ağaçları, meşe ağaçları ve paçavra otları gibi rüzgarla dağılan tozlayıcılar tarafından üretilen polenlerdir; polenlerdeki alerjenler tozlaşma sürecinde gerekli olduğu düşünülen proteinlerdir. ⓘ
Döllenme
_(1914)_(14590686978).jpg)
Sinagmi olarak da adlandırılan döllenme, polenlerin ercikten karpele hareketi olan tozlaşmayı takiben gerçekleşir. Hem protoplastların birleşmesi olan plazmogamiyi hem de çekirdeklerin birleşmesi olan karyogamiyi kapsar. Polen çiçeğin stigması üzerine düştüğünde, stilden aşağıya ve yumurtalığa doğru uzanan bir polen tüpü oluşturmaya başlar. Yumurtalığın en orta kısmına girdikten sonra yumurta aparatına ve bir sinerjide girer. Bu noktada polen tüpünün ucu patlar ve iki sperm hücresini serbest bırakır, bunlardan biri hücre zarını ve protoplazmasının çoğunu kaybederken yumurtaya doğru yol alır. Sperm çekirdeği daha sonra yumurta çekirdeği ile birleşerek diploid (her kromozomdan iki kopya) bir hücre olan zigotun oluşmasını sağlar. ⓘ
Döllenmede sadece plazmogami ya da tüm eşey hücrelerinin kaynaşması gerçekleşirken, Angiospermlerde (çiçekli bitkiler) hem karyogami hem de plazmogamiyi içeren çift döllenme olarak bilinen bir süreç gerçekleşir. Çifte döllenmede ikinci sperm hücresi de daha sonra sinerjitin içine girer ve merkezi hücrenin iki kutup çekirdeği ile birleşir. Her üç çekirdek de haploid olduğundan, triploid olan büyük bir endosperm çekirdeği ile sonuçlanırlar. ⓘ
Tohum gelişimi

Zigot oluşumunu takiben mitoz adı verilen nükleer ve hücresel bölünmeler yoluyla büyümeye başlar ve sonunda küçük bir hücre grubu haline gelir. Bir bölümü embriyo olurken, diğer bölümü embriyoyu endospermin içine iten ve daha sonra fark edilemeyen bir yapı olan suspensör haline gelir. Bu sırada daha sonra enerji deposu olarak kullanılan kotiledon haline gelecek olan iki küçük primordia da oluşur. Bu primordialardan bir tanesini büyüten bitkilere monokotiledon, ikisini büyütenlere ise dikotiledon adı verilir. Bir sonraki aşama Torpido aşaması olarak adlandırılır ve radikül (embriyotik kök), epikotil (embriyotik gövde) ve hipokotil (kök/sürgün birleşimi) gibi birkaç temel yapının büyümesini içerir. Son aşamada tohumun etrafında vasküler doku gelişir. ⓘ
Meyve gelişimi
İçinde ovülden tohumun oluştuğu ovaryum büyüyerek meyveye dönüşür. Diğer tüm ana çiçek parçaları bu gelişim sırasında ölür: stil, stigma, çanak yapraklar, stamenler ve taç yapraklar. Meyve üç yapı içerir: ekzokarp veya dış tabaka, mezokarp veya etli kısım ve endokarp veya en iç tabaka, meyve duvarı ise perikarp olarak adlandırılır. Boyut, şekil, sertlik ve kalınlık farklı meyveler arasında değişiklik gösterir. Bunun nedeni, tohumun dağılma yöntemiyle doğrudan bağlantılı olmasıdır; meyvenin amacı budur - tohumun dağılmasını teşvik etmek veya sağlamak ve bunu yaparken tohumu korumak. ⓘ
Tohumun dağılması
.jpg)


Bir çiçeğin tozlaşmasını, döllenmeyi ve nihayetinde bir tohum ve meyvenin gelişimini takiben, meyveyi bitkiden uzaklaştırmak için tipik olarak bir mekanizma kullanılır. Kapalı tohumlu bitkilerde (çiçekli bitkiler) tohumlar, ana ve yavru bitkiler arasında rekabete neden olmamak ve yeni alanların kolonileşmesini sağlamak için bitkiden uzağa dağılır. Genellikle iki kategoriye ayrılırlar, ancak birçok bitki bunların arasında veya bir ya da daha fazlasında yer alır: ⓘ
Allochory
Allokoride, bitkiler tohumlarını kendilerinden uzağa taşımak için harici bir vektör ya da taşıyıcı kullanırlar. Bunlar kuşlar ve karıncalar gibi biyotik (canlı) ya da rüzgar veya su gibi abiyotik (cansız) olabilir. ⓘ
Biyotik vektörler
Birçok bitki, tohumlarını kendilerinden uzağa dağıtmak için biyotik vektörler kullanır. Bu yöntem Zoochory şemsiye terimine girerken, fruigivory olarak da bilinen Endozoochory, özellikle hayvanları onları yemeye çekmek için meyve yetiştirmeye adapte olmuş bitkileri ifade eder. Yenildikten sonra tipik olarak hayvanın sindirim sisteminden geçerler ve bitkiden uzağa dağılırlar. Bazı tohumlar ya hayvanların taşlığında dayanacak şekilde ya da hatta hayvanlardan geçtikten sonra daha iyi çimlenecek şekilde özel olarak uyarlanmıştır. Kuşlar (ornithochory), yarasalar (chiropterochory), kemirgenler, primatlar, karıncalar (myrmecochory), kuş olmayan sauropsidler (saurochory), genel olarak memeliler (mammaliochory) ve hatta balıklar tarafından yenebilirler. Tipik olarak meyveleri etlidir, yüksek besin değerine sahiptir ve dağıtıcılar için ek bir "ödül" olarak kimyasal cezbedicilere sahip olabilir. Bu durum morfolojik olarak daha fazla posa, bir aril ve bazen de diğer etli yapılar olan elaiosome (özellikle karıncalar için) varlığında kendini gösterir. Epizoochory, Acaena cinsindeki birçok tür gibi tohumları hayvanlara tutunmaya ve bu şekilde dağılmaya adapte olmuş bitkilerde görülür. Tipik olarak bu bitkilerin tohumları, kuşlar ve kürklü hayvanlar da dahil olmak üzere hayvanlara daha kolay tutunmak için kancalara veya yapışkan bir yüzeye sahiptir. Bazı bitkiler hayvanları kandırarak tohumlarını dağıtmak için mimesis ya da taklit yöntemini kullanır ve bu bitkiler genellikle özel olarak uyarlanmış renklere sahiptir. Zoochory'nin son türü Synzoochory olarak adlandırılır; bu türde ne tohumların sindirilmesi ne de tohumların vücutta kasıtsız olarak taşınması söz konusudur, ancak tohumlar hayvanlar tarafından kasıtlı olarak taşınır. Bu genellikle hayvanın ağzında ya da gagasında gerçekleşir (Stomatochory olarak adlandırılır), birçok kuş ve tüm karıncalar için kullanılan da budur.

.jpg)
Abiyotik vektörler
Abiyotik dağılımda bitkiler tohumlarını kendilerinden uzağa taşımak için rüzgar, su ya da kendilerine ait bir mekanizmanın vektörlerini kullanırlar. Anemokori, bitki tohumlarını dağıtmak için rüzgarı bir vektör olarak kullanmayı içerir. Bu tohumlar rüzgarda hareket etmek zorunda olduklarından neredeyse her zaman küçüktürler - hatta bazen toz gibidirler, yüksek yüzey alanı/hacim oranına sahiptirler ve çok sayıda üretilirler - bazen bir milyona kadar. Takla otu gibi bitkiler, tohumların rüzgarla birlikte yuvarlanmasına izin vermek için tüm sürgünü ayırır. Bir başka yaygın adaptasyon da tohumların havada daha uzun süre kalmasını ve dolayısıyla daha uzağa gitmesini sağlayan kanatlar, tüyler veya balon benzeri yapılardır. Hidrokoryada bitkiler tohumlarını su kütleleri aracılığıyla dağıtmaya adapte olmuşlardır ve bu nedenle tipik olarak kaldırma kuvvetine ve suya göre düşük bir bağıl yoğunluğa sahiptirler. Genellikle tohumlar morfolojik olarak hidrofobik yüzeyler, küçük boyut, tüyler, balçık, yağ ve bazen tohumların içinde hava boşlukları ile uyarlanmıştır. Bu bitkiler üç kategoriye ayrılır: tohumların su akıntılarının yüzeyinde, su akıntılarının yüzeyinin altında ve yağmurun bir bitkiye inmesiyle dağıldığı bitkiler. ⓘ
Otokori
Otokoride, bitkiler tohumları kendilerinden uzaklaştırmak için kendi vektörlerini yaratırlar. Bunun için adaptasyonlar genellikle meyvelerin patlamasını ve tohumları Hura crepitans'ta olduğu gibi balistik olarak uzağa itmesini veya bazen sürünen diasporların yaratılmasını içerir. Bu yöntemlerin tohumlarını dağıtabileceği mesafeler nispeten küçük olduğundan, genellikle harici bir vektörle eşleştirilirler. ⓘ
Evrim
Kara bitkileri yaklaşık 425 milyon yıldır var olsa da, ilk bitkiler suda yaşayan benzerlerinin basit bir uyarlamasıyla ürediler: sporlar. Denizde bitkiler -ve bazı hayvanlar- kendilerinin genetik klonlarını yüzüp başka bir yerde büyümeleri için basitçe saçabilirler. İlk bitkiler bu şekilde çoğaldı. Ancak bitkiler kısa süre sonra, karada denizdekinden daha olası olan kuruma ve diğer hasarlarla başa çıkmak için bu kopyaları koruma yöntemleri geliştirdi. Henüz çiçek evrimleşmemiş olsa da, koruma tohum haline geldi. İlk tohum taşıyan bitkiler arasında ginkgo ve kozalaklı ağaçlar bulunmaktadır. ⓘ

Başta tohum eğrelti otları olmak üzere soyu tükenmiş birkaç açık tohumlu bitki grubunun çiçekli bitkilerin atası olduğu öne sürülmüştür, ancak çiçeklerin tam olarak nasıl evrimleştiğini gösteren sürekli bir fosil kanıtı yoktur. Fosil kayıtlarında nispeten modern çiçeklerin aniden ortaya çıkması evrim teorisi için öyle bir sorun teşkil etmiştir ki Charles Darwin tarafından "iğrenç bir gizem" olarak adlandırılmıştır. ⓘ
Yakın zamanda keşfedilen Archaefructus gibi anjiyosperm fosilleri ve yeni keşfedilen gymnosperm fosilleri, anjiyosperm özelliklerinin bir dizi adımda nasıl kazanılmış olabileceğini düşündürmektedir. Çiçekli bir bitkinin erken fosili olan Çin'den Archaefructus liaoningensis, yaklaşık 125 milyon yıl öncesine tarihlenmektedir. Çin'den daha da eski olan 125-130 milyon yıllık Archaefructus sinensis'tir. 2015 yılında İspanya'da keşfedilen bir bitkinin (130 milyon yaşındaki Montsechia vidalii) 130 milyon yaşında olduğu iddia edilmiştir. 2018'de bilim insanları, en erken çiçeklerin yaklaşık 180 milyon yıl önce başladığını bildirdi. ⓘ
_fragment.jpg)
Yakın zamanda yapılan DNA analizleri (moleküler sistematik), Pasifik'teki Yeni Kaledonya adasında bulunan Amborella trichopoda'nın çiçekli bitkilerin geri kalanıyla kardeş grupta yer alan tek tür olduğunu ve morfolojik çalışmaların da bu türün en eski çiçekli bitkilerin karakteristik özelliklerini taşıdığını gösterdiğini ortaya koymuştur. ⓘ
Kretase döneminde ya da hemen öncesinde çiçeklerin varlığına dair kesin kanıtların yanı sıra, 250 milyon yıl kadar önce çiçeklerin varlığına dair bazı ikinci derece kanıtlar da bulunmaktadır. Bitkilerin çiçeklerini savunmak için kullandığı bir kimyasal olan oleanane, o dönemde evrimleşen ve modern, çiçekli bitkilerin birçok özelliğini taşıyan gigantopteridler de dahil olmak üzere o kadar eski fosil bitkilerde tespit edilmiştir, ancak bunların çiçekli bitkiler olduğu bilinmemektedir, çünkü sadece gövdeleri ve dikenleri ayrıntılı olarak korunmuş olarak bulunmuştur; taşlaşmanın en eski örneklerinden biri. ⓘ
Yaprak ve gövde yapısındaki benzerlik çok önemli olabilir, çünkü çiçekler genetik olarak bitkilerdeki normal yaprak ve gövde bileşenlerinin bir adaptasyonudur ve normalde yeni sürgünler oluşturmaktan sorumlu genlerin bir kombinasyonudur. En ilkel çiçeklerin, genellikle birbirlerinden ayrı (ancak temas halinde) değişken sayıda çiçek parçasına sahip olduğu düşünülmektedir. Çiçekler sarmal bir şekilde büyüme, biseksüel olma (bitkilerde bu, aynı çiçekte hem erkek hem de dişi kısımlar anlamına gelir) ve yumurtalık (dişi kısım) tarafından domine edilme eğiliminde olurdu. Çiçekler daha da geliştikçe, bazı varyasyonlar, çok daha spesifik bir sayı ve tasarımla ve çiçek veya bitki başına belirli cinsiyetlerle veya en azından "alt yumurtalık" ile birbirine kaynaşmış parçalar geliştirmiştir. ⓘ
Genel varsayım, çiçeklerin işlevinin en başından beri hayvanları üreme sürecine dahil etmek olduğu yönündedir. Polenler, parlak renkler ve belirgin şekiller olmadan saçılabilir, bu nedenle başka bir fayda sağlamadıkları sürece bitkinin kaynaklarını kullanmak bir sorumluluk olacaktır. Çiçeklerin aniden, tamamen gelişmiş bir şekilde ortaya çıkmasının önerilen bir nedeni, bir ada veya adalar zinciri gibi izole bir ortamda evrimleşmeleri ve onları taşıyan bitkilerin belirli bir hayvanla (örneğin bir yaban arısı), bugün birçok ada türünün geliştiği gibi, son derece uzmanlaşmış bir ilişki geliştirebilmeleridir. Bugün incir eşek arılarının yaptığı gibi bir bitkiden diğerine polen taşıyan varsayımsal bir eşek arısı ile bu simbiyotik ilişki, sonunda hem bitki(ler)in hem de ortaklarının yüksek derecede uzmanlaşma geliştirmesiyle sonuçlanmış olabilir. Ada genetiğinin, özellikle de daha düşük geçiş formları gerektirmiş gibi görünen radikal adaptasyonlar söz konusu olduğunda, türleşmenin yaygın bir kaynağı olduğuna inanılmaktadır. Yaban arısı örneğinin tesadüfi olmadığına dikkat edin; görünüşe göre özellikle simbiyotik bitki ilişkileri için evrimleşmiş olan arılar, yaban arılarının soyundan gelmektedir. ⓘ
Benzer şekilde, bitki üremesinde kullanılan meyvelerin çoğu çiçeğin bazı kısımlarının büyütülmesiyle elde edilir. Bu meyve sıklıkla hayvanların onu yemek istemesine ve böylece içerdiği tohumları saçmasına bağlı olan bir araçtır. ⓘ
Bu tür simbiyotik ilişkilerin birçoğu anakaradaki organizmalarla rekabette hayatta kalamayacak kadar kırılgan olsa da, çiçekler alışılmadık derecede etkili bir üretim aracı olduklarını kanıtlamış ve (asıl kökenleri ne olursa olsun) yayılarak kara bitki yaşamının baskın biçimi haline gelmiştir. ⓘ
Çiçek evrimi günümüze kadar devam etmektedir; modern çiçekler insanlar tarafından o kadar derinden etkilenmiştir ki birçoğu doğada tozlaşamaz. Modern, evcilleştirilmiş çiçeklerin çoğu eskiden sadece toprak bozulduğunda filizlenen basit yabani otlardı. Bazıları insan mahsulleriyle birlikte büyüme eğilimindeydi ve en güzelleri güzellikleri nedeniyle koparılmadı, insan sevgisine bağımlı ve özel bir adaptasyon geliştirdi. ⓘ
Renk
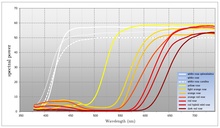
Birçok çiçekli bitki, bitkinin çekmeyi amaçladığı tozlaştırıcının görünür dalga boyları aralığında mümkün olduğunca fazla ışığı yansıtır. Görünür ışık aralığının tamamını yansıtan çiçekler genellikle bir insan gözlemci tarafından beyaz olarak algılanır. Beyaz çiçeklerin önemli bir özelliği, görünür spektrumun tamamını eşit şekilde yansıtmalarıdır. Birçok çiçekli bitki tozlayıcıları çekmek için beyazı kullanırken, renk kullanımı da yaygındır (aynı tür içinde bile). Renk, çiçekli bir bitkinin çekmek istediği tozlayıcı hakkında daha spesifik olmasını sağlar. İnsan renk üretim teknolojisi (CMYK) tarafından kullanılan renk modeli, spektrumu geniş emilim alanlarına bölen pigmentlerin modülasyonuna dayanır. Buna karşın çiçekli bitkiler emilim ve yansıma arasındaki geçiş noktası dalga boyunu kaydırabilmektedir. Çoğu tozlaştırıcının görsel sistemlerinin görünür spektrumu dairesel olarak gördüğü varsayılırsa, çiçekli bitkilerin spektrumun bir bölgesindeki ışığı emerek ve diğer bölgedeki ışığı yansıtarak renk ürettiği söylenebilir. CMYK ile renk, geniş emilim bölgelerinin genliğinin bir fonksiyonu olarak üretilir. Buna karşın çiçekli bitkiler, yansıtılan ışığın frekansını (ya da daha doğrusu dalga boyunu) değiştirerek renk üretir. Çoğu çiçek spektrumun maviden sarıya kadar olan bölgesindeki ışığı emer ve spektrumun yeşilden kırmızıya kadar olan bölgesindeki ışığı yansıtır. Birçok çiçekli bitki türü için, ürettikleri rengi karakterize eden geçiş noktasıdır. Renk, soğurma ve yansıtma arasındaki geçiş noktası kaydırılarak modüle edilebilir ve bu şekilde çiçekli bir bitki hangi tozlaştırıcıyı çekmek istediğini belirleyebilir. Bazı çiçekli bitkiler de emilim alanlarını modüle etme konusunda sınırlı bir yeteneğe sahiptir. Bu genellikle dalga boyu üzerindeki kontrol kadar kesin değildir. İnsan gözlemciler bunu doygunluk derecesi (renkteki beyaz miktarı) olarak algılayacaktır. ⓘ
Klasik taksonomi
.png)
Bitkilerin sınıflandırılması ve tanımlanması çalışması olan bitki taksonomisinde, bitki çiçeklerinin morfolojisi yaygın olarak kullanılmaktadır ve binlerce yıldır kullanılmaktadır. Bitki taksonomisinin tarihi Theophrastus'un yazılarıyla en azından M.Ö. 300'lere kadar uzansa da, modern bilimin temeli 18. ve 19. yüzyıllardaki çalışmalara dayanmaktadır. ⓘ
Carl Linnaeus (1707-1778), çalışma hayatının çoğunu doğa tarihi profesörü olarak geçirmiş İsveçli bir botanikçidir. Dönüm noktası niteliğindeki 1757 tarihli Species Plantarum adlı kitabı, sınıflandırma sisteminin yanı sıra bugün hala dünya çapında kullanılan binomial isimlendirme kavramını da ortaya koymaktadır. Temel olarak stamenlerin sayısı, uzunluğu ve birleşimine dayanan 24 sınıf tanımlamıştır. İlk on sınıf doğrudan stamen sayısını takip ederken (Octandria 8 stamene sahiptir vb.), on birinci sınıf 11-20 stamene ve on iki ve on üçüncü sınıflar 20 stamene sahiptir; sadece bağlanma noktalarında farklılık gösterirler. Sonraki beş sınıf stamenlerin uzunluğuyla, son beş sınıf ise bitkinin üreme kabiliyetinin doğasıyla, stamenlerin nerede büyüdüğüyle ve çiçeğin gizlenip gizlenmediğiyle (eğrelti otlarında olduğu gibi) ilgilidir. Yapay olmasına rağmen bu sınıflandırma yöntemi, yerini başka bir botanikçinin sistemine bırakmadan önce, takip eden yetmiş yıl boyunca yaygın olarak kullanılmıştır. ⓘ
Antoine Laurent de Jussieu (1748-1836), 1787 tarihli Genera plantarum: secundum ordines naturales disposita adlı çalışmasıyla bitkileri sınıflandırmak için yeni bir yöntem ortaya koyan Fransız bir botanikçiydi; bunun yerine doğal özelliklere dayanıyordu. Bitkiler, varsa kotiledon sayısına ve stamenlerin konumuna göre ayrılıyordu. Bir sonraki en önemli sınıflandırma sistemi 19. yüzyılın sonlarında botanikçiler Joseph Dalton Hooker (1817-1911) ve George Bentham'dan (1800-1884) geldi. De Jussieu ve Augustin Pyramus de Candolle'un daha önceki çalışmaları üzerine inşa ettiler ve hala dünyanın birçok herbarisinde kullanılan bir sistem geliştirdiler. Bitkiler en üst düzeyde kotiledon sayısına ve çiçeklerin doğasına göre ayrılmış, daha sonra takımlara (familyalara), cinslere ve türlere ayrılmıştır. Bu sınıflandırma sistemi 1862 ve 1883 yılları arasında Genera plantarum adlı üç cilt halinde yayımlanmıştır. En çok kabul gören ve bazı ortamlarda "en iyi sınıflandırma sistemi" olarak kabul edilen sistemdir. ⓘ
Darwin'in Türlerin Kökeni kitabından sonra bilimsel düşüncede yaşanan gelişmenin ardından, birçok botanikçi daha filogenetik yöntemler kullanmış ve genetik dizileme, sitoloji ve palinoloji kullanımı giderek yaygınlaşmıştır. Buna rağmen, çiçek ve çiçek salkımının doğası gibi morfolojik özellikler hala bitki taksonomisinin temelini oluşturmaktadır. ⓘ
Sembolizm

Batı kültüründe birçok çiçeğin önemli sembolik anlamları vardır. Çiçeklere anlamlar yükleme uygulaması floriografi olarak bilinir. En yaygın örneklerden bazıları şunlardır:
- Kırmızı güller aşkın, güzelliğin ve tutkunun sembolü olarak verilir.
- Gelincikler ölüm zamanında tesellinin sembolüdür. Birleşik Krallık, Yeni Zelanda, Avustralya ve Kanada'da savaş zamanlarında ölen askerleri anmak için kırmızı gelincikler takılır.
- Süsen/Zambak "diriliş/yaşam" anlamına gelen bir sembol olarak cenaze törenlerinde kullanılır. Ayrıca yıldızlarla (güneş) ve çiçek açan/parlayan yapraklarıyla da ilişkilendirilir.
- Papatyalar masumiyetin sembolüdür. ⓘ

Çiçekler, çeşitli ve renkli görünümleri nedeniyle uzun zamandır görsel sanatçıların da favori konusu olmuştur. Van Gogh'un ayçiçekleri serisi veya Monet'nin nilüferleri gibi tanınmış ressamların en ünlü tablolarından bazıları çiçeklerden oluşmaktadır. Çiçekler ayrıca kalıcı, üç boyutlu çiçek sanatı parçaları yaratmak için kurutulur, dondurularak kurutulur ve preslenir. ⓘ
Sanatta çiçekler, Georgia O'Keeffe, Imogen Cunningham, Veronica Ruiz de Velasco ve Judy Chicago gibi sanatçıların eserlerinde ve aslında Asya ve batı klasik sanatında görüldüğü gibi kadın cinsel organını da temsil eder. Dünyadaki pek çok kültürde çiçekleri kadınsılıkla ilişkilendirme eğilimi belirgindir. ⓘ
Narin ve güzel çiçeklerin büyük çeşitliliği, özellikle 18-19. yüzyıl Romantik döneminden çok sayıda şairin eserlerine ilham vermiştir. Ünlü örnekler arasında William Wordsworth'un I Wandered Lonely as a Cloud ve William Blake'in Ah! Sun-Flower adlı şiirleri sayılabilir. ⓘ
Rüyalardaki sembolizmi de "çiçek açma potansiyeli" gibi olası yorumlarla tartışılmıştır. ⓘ
Çiçeklerin, bahçelerin ve bahar mevsiminin Roma tanrıçası Flora'dır. Yunan bahar, çiçek ve doğa tanrıçası Chloris'tir. ⓘ
Hindu mitolojisinde çiçekler önemli bir yere sahiptir. Hindu sistemindeki üç büyük tanrıdan biri olan Vişnu, genellikle bir lotus çiçeğinin üzerinde dimdik dururken tasvir edilir. Vişnu ile olan ilişkisinin yanı sıra, Hindu geleneği lotusun ruhani bir öneme sahip olduğunu da düşünür. Örneğin, Hindu yaratılış hikayelerinde yer alır. ⓘ
İnsan kullanımı

Tarih, çiçeklerin insanlar tarafından binlerce yıldır çeşitli amaçlara hizmet etmek için kullanıldığını göstermektedir. Bunun erken bir örneği, çiçeklerin kadınların saçlarını süslemek için kullanıldığı Antik Mısır'da yaklaşık 4.500 yıl öncesine aittir. Çiçekler aynı zamanda Monet'nin Nilüferler tablosunda ya da William Wordsworth'ün nergisler hakkında yazdığı "Bir Bulut Kadar Yalnız Dolaştım" adlı şiirinde olduğu gibi sanata da defalarca ilham vermiştir. ⓘ
Modern zamanlarda insanlar, kısmen hoş görünümleri ve kokuları nedeniyle çiçek ve çiçek açan bitkiler yetiştirmenin, satın almanın, giymenin veya başka şekillerde etraflarında olmanın yollarını aramışlardır. Dünyanın her yerinde insanlar çiçekleri hayatlarındaki önemli olayları işaretlemek için kullanırlar:
- Yeni doğumlar veya vaftizler için
- Sosyal etkinliklerde veya tatillerde takılan bir korsaj veya yaka çiçeği olarak
- Sevgi ya da saygı göstergesi olarak
- Gelin partisi için düğün çiçekleri ve salon süslemeleri için
- Ev içinde aydınlatıcı süslemeler olarak
- İyi yolculuklar partileri, eve hoş geldin partileri ve "seni düşünüyorum" hediyeleri için bir hatıra hediyesi olarak
- Cenaze çiçekleri ve yas tutanlar için sempati ifadeleri için
- İbadet için. Hıristiyanlıkta kilise çiçekleri genellikle kiliseleri süsler. Hindu kültüründe, taraftarlar genellikle tapınaklara hediye olarak çiçek getirir ⓘ

Yasemin gibi çiçekler yüzyıllardır Çin'de geleneksel çayın yerine kullanılmaktadır. Son zamanlarda dünya çapında geleneksel olarak kullanılan diğer birçok bitki ve çiçek, bir dizi Çiçek Çayı hazırlamak için önem kazanmaktadır. ⓘ
Bu nedenle insanlar evlerinin çevresinde çiçek yetiştirmekte, yaşam alanlarının bir bölümünü çiçek bahçelerine ayırmakta, kır çiçekleri toplamakta ya da çiçekçilerden ticari olarak yetiştirilen çiçekleri satın almaktadır. ⓘ

Çiçekler diğer ana bitki kısımlarından (tohumlar, meyveler, kökler, saplar ve yapraklar) daha az gıda sağlar, ancak yine de birkaç önemli sebze ve baharat sağlar. Çiçek sebzeleri arasında brokoli, karnabahar ve enginar bulunmaktadır. En pahalı baharat olan safran, çiğdemin kurutulmuş stigmalarından oluşur. Diğer çiçek baharatları karanfil ve kaparidir. Şerbetçiotu çiçekleri birayı tatlandırmak için kullanılır. Kadife çiçeği çiçekleri tavuklara yedirilerek yumurta sarılarına tüketicilerin daha çok arzu ettiği altın sarısı renk verilir; kurutulmuş ve öğütülmüş kadife çiçeği çiçekleri de Gürcü mutfağında baharat ve renklendirici olarak kullanılır. Karahindiba ve mürver çiçekleri genellikle şarap haline getirilir. Arı poleni, arılardan toplanan polen, bazı insanlar tarafından sağlıklı bir gıda olarak kabul edilir. Bal, arıların işlediği çiçek nektarından oluşur ve genellikle çiçeğin türüne göre adlandırılır; örneğin portakal çiçeği balı, yonca balı ve tupelo balı. ⓘ
Yüzlerce taze çiçek yenilebilir, ancak sadece birkaçı yaygın olarak gıda olarak pazarlanmaktadır. Genellikle salatalara garnitür olarak eklenirler. Kabak çiçekleri ekmek kırıntılarına batırılır ve kızartılır. Bazı yenilebilir çiçekler arasında nasturtium, krizantem, karanfil, kedi kuyruğu, Japon hanımeli, hindiba, peygamber çiçeği, kanna ve ayçiçeği bulunur. Papatya, gül ve menekşe gibi yenilebilir çiçekler bazen şekerlenir. ⓘ
Kokuları ve tıbbi özellikleri nedeniyle seçilen krizantem, gül, yasemin, Japon hanımeli ve papatya gibi çiçekler çayla karıştırılarak ya da tek başlarına tisan olarak kullanılır. ⓘ
Çiçekler tarih öncesi çağlardan beri cenaze törenlerinde kullanılmaktadır: İspanya'daki El Miron Mağarası'nda bulunan bir kadın mezarında polen izlerine rastlanmıştır. Birçok kültür çiçekler ile yaşam ve ölüm arasında bir bağlantı kurar ve mevsimsel dönüşleri nedeniyle çiçekler yeniden doğuşu da çağrıştırır, bu da birçok insanın neden mezarlara çiçek koyduğunu açıklayabilir. Euripides'in Fenikeli Kadınlar adlı oyununda kaydedildiği gibi, eski Yunanlılar ölen kişinin başına çiçeklerden bir taç yerleştirir; ayrıca mezarları çelenkler ve çiçek yapraklarıyla kaplarlardı. Eski Mısır mezarlarında çiçekler yaygın olarak kullanılıyordu ve Meksikalılar bugün de Aztek atalarının yaptığı gibi Ölüler Günü kutlamalarında çiçekleri belirgin bir şekilde kullanmaktadır. ⓘ

Vermek

Çiçek verme geleneği, çiçeklerin genellikle tıbbi ve bitkisel niteliklere sahip olduğu tarih öncesi dönemlere kadar uzanmaktadır. Arkeologlar birçok mezar alanında çiçek yapraklarının kalıntılarını bulmuşlardır. Çiçekler ilk olarak kurban ve gömü nesnesi olarak kullanılmıştır. Eski Mısırlılar ve daha sonra Yunanlılar ve Romalılar çiçekleri kullanmışlardır. Mısır'da, M.Ö. 1540 yıllarına ait, kırmızı gelincik, sarı Araun, peygamber çiçeği ve zambak tasvirli gömü nesneleri bulunmuştur. Çiçek verme kayıtları Çin yazılarında ve Mısır hiyerogliflerinin yanı sıra Yunan ve Roma mitolojisinde de yer almaktadır. Çiçek verme uygulaması, çiftlerin sevgilerini çiçekler aracılığıyla gösterdikleri Orta Çağ'da gelişmiştir. ⓘ
Çiçek verme geleneği birçok biçimde varlığını sürdürmektedir. Rus kültürünün ve folklorunun önemli bir parçasıdır. Öğrencilerin öğretmenlerine çiçek vermesi yaygındır. Romantik bir ilişkide sarı çiçek vermek Rusya'da ayrılık anlamına gelir. Günümüzde çiçekler genellikle bir çiçek buketi şeklinde verilmektedir. ⓘ
Çanak yapraklar (sepal)
Çiçeğin yeşil veya kahverengi tonlarında olan minik yapraklardır. Bu yapraklar, çiçek tomurcuk halindeyken onu dış etkenlerden korur. Ancak çiçeğin eşi uyumsuz ise çanak yaprak diye bir şey yoktur. ⓘ
Taç yapraklar (petal)
Çiçeğin en gösterişli kısmıdır. Üreme organlarının dışında bulunur. Gösterişli yapısı ve renkleriyle birçok canlıyı cezbederek tozlaşmada önemli bir role sahiptir. İçindeki kromoplast organeli ise yeşil hariç diğer renkleri salgılar. Bu da tozlaşmada önemli rol oynar. ⓘ
Monokotil bitkilerde çiçek örtüsü 'tepal' adı verilen bir kısımdan oluşur. Taç yapraklar gösterişli, güzel kokan ve renkli olduklarından böcekleri kendilerine çeker ve bu olay sonucunda arılar veya böcekler polen depo etmiş olurlar. ⓘ
Üreme organları
Üreme işlevini yerine getirerek neslin devamlılığını sağlar. Bu nedenle 'verimli kısım' olarak da adlandırılır. ⓘ
Erkek üreme organı
Filament (sapçık) ve anter (başçık) olmak üzere 2 kısımdan oluşur. Filament, anterin çiçeğe tutmasını; anter ise, polen oluşumunu sağlar. ⓘ